Nous recrutons ! Rejoignez-nous et faites la différence au sein de nos hôpitaux
Enseignement pour les professionnels
L’excellence et la qualité des soins passent aussi par la formation des futurs soignants.
L’HUDERF, un Hôpital Universitaire
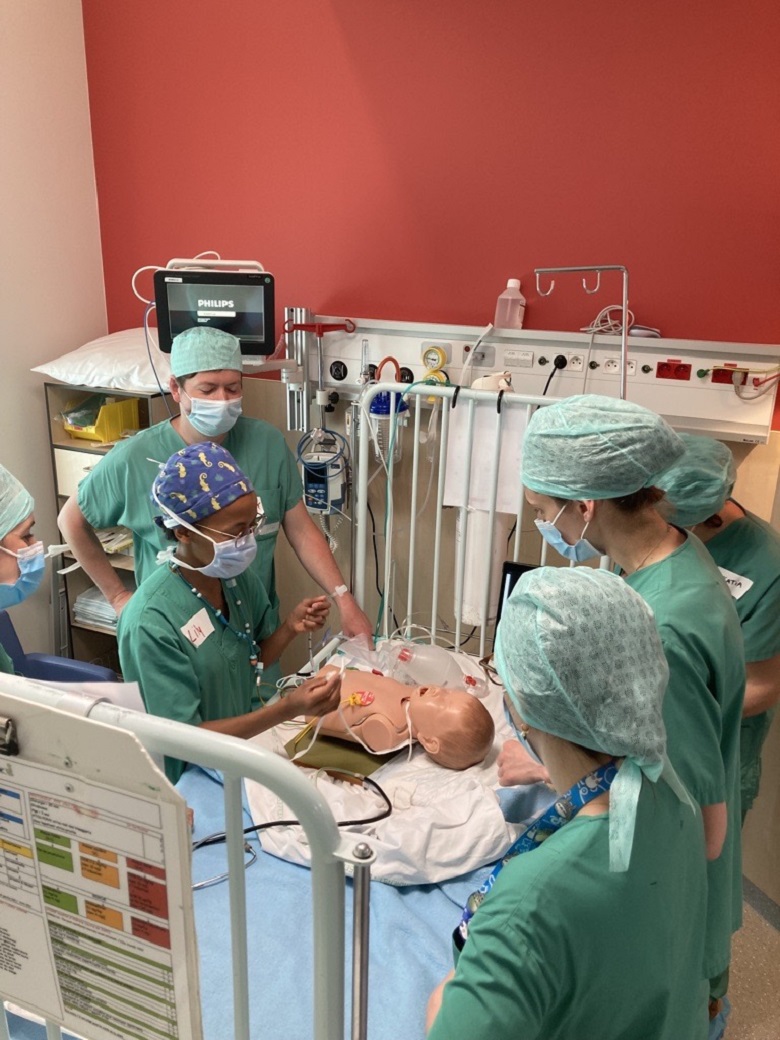
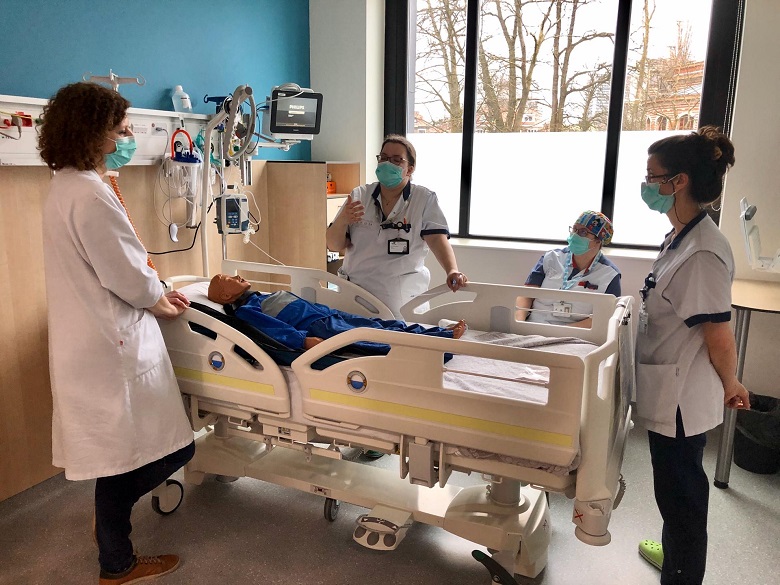
Le rôle de l'HUDERF, en sa qualité d’hôpital universitaire, est aussi de contribuer activement à la transmission des connaissances médicales. Nos équipes œuvrent à enrichir et à perfectionner les soins aux patients par un enseignement de qualité. Les méthodes innovantes d’enseignement permettent à toutes les professions de santé d’acquérir les compétences nécessaires avant de se plonger dans l’apprentissage sur le terrain. L’enseignement se déroule également sur le terrain, quotidiennement, au plus proche des équipes médicales et des patients. L’enseignement est dirigé non seulement vers les générations futures de soignants mais également vers les collaborateurs actuels au travers de nombreuses formations proposées.
En termes de prise en charge, le respect du choix du patient reste une valeur fondamentale dans le cadre de nos activités d’enseignement.
Vous êtes notre prochain collaborateur ?
Travailler à l’Hôpital des Enfants pendant ou juste après ses études est une occasion unique de développer ses compétences et de faire connaissance avec un environnement de travail et de soins tout à fait unique.
Plus d’informations sur les stages :


{kind=link}
{kind=link}