We zijn aan het werven ! Kom bij ons en maak het verschil in onze ziekenhuizen
Onderwijs voor professionals
Uitmuntendheid en kwaliteit van de zorgverlening hebben ook te maken met de opleiding van de toekomstige zorgverleners.
HUDERF, een universitair ziekenhuis
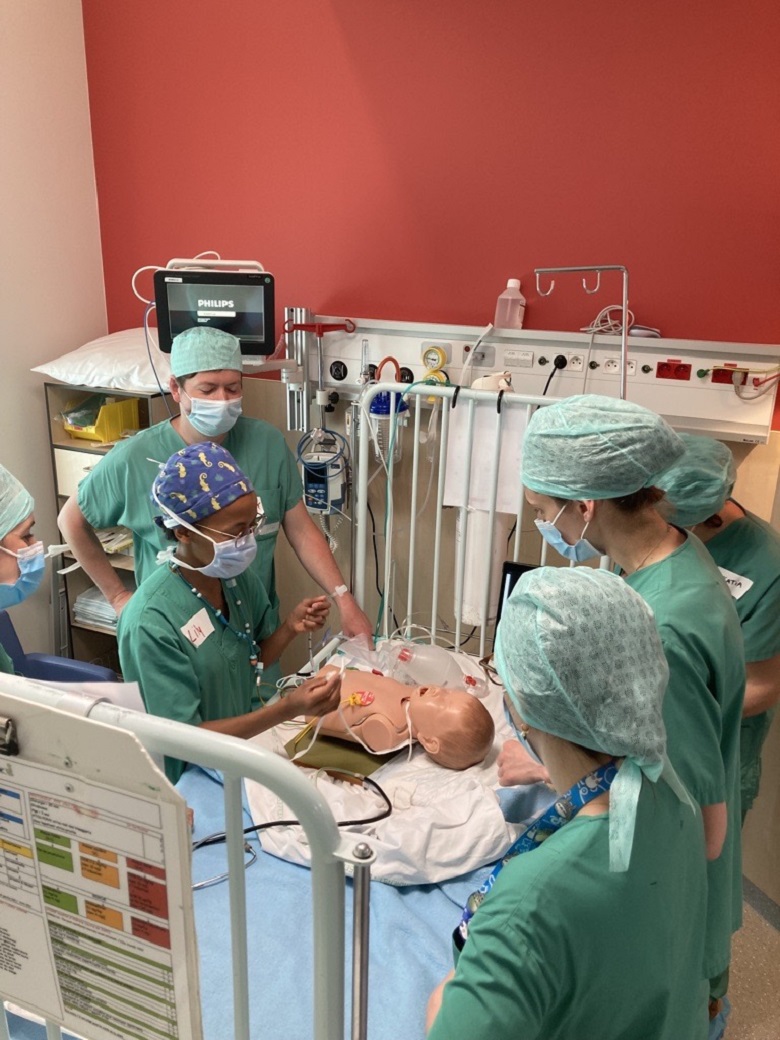
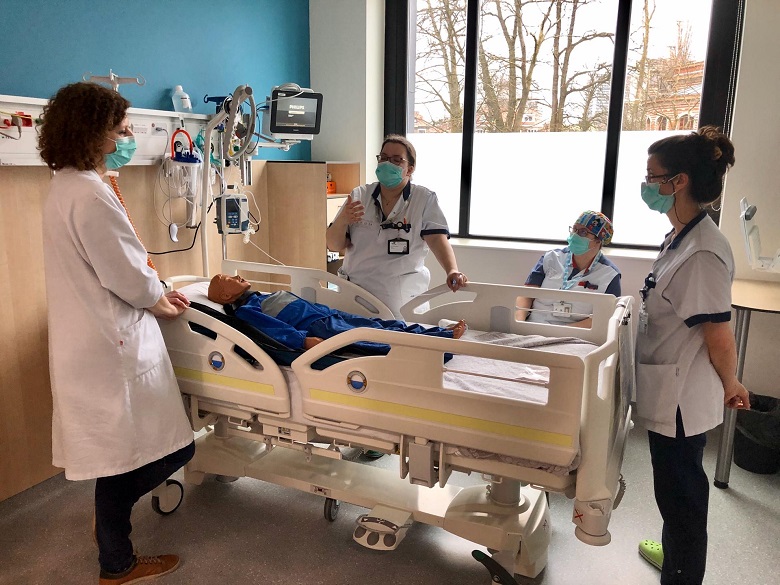
Een belangrijke rol van HUDERF - als academisch ziekenhuis - is actief bij te dragen tot de overdracht van medische kennis. Onze teams spannen zich in om de zorgverlening aan patiënten te verrijken en te perfectioneren dankzij kwaliteitsvol onderwijs. De innoverende onderwijsmethodes maken het voor alle beroepen in de gezondheidssector mogelijk om de nodige competenties te verwerven vooraleer zich onder te dompelen in leren op de werkvloer. Het onderwijs speelt zich ook op het terrein af, dagelijks, zo dicht mogelijk bij de medische teams en de patiënten. Het onderwijs is niet alleen gericht op de komende generaties zorgverleners maar ook op de huidige medewerkers, via de vele aangeboden opleidingen.
Wat benadering van de patiënt aangaat, blijft respect voor de keuze van de patiënt een fundamentele waarde in het kader van onze onderwijsactiviteiten.
Word jij onze nieuwe medewerker?
Werken in het Kinderziekenhuis, tijdens of meteen na je studies, is een unieke kans om je competenties te ontwikkelen en om kennis te maken met een absoluut unieke werk- en zorgomgeving.
Meer informatie over stages :


{kind=link}
{kind=link}